







"LIBRAS"
(LÍNGUA BRASILEIRA DE SINAIS)
UM ESTUDO ELETROENCEFALOGRAFICO DE SUA FUNCIONALIDADE CEREBRAL
Fábio Theoto Rocha
Gerente da EINA - Estudos em Inteligência Natural e Artificial Ltda
Introdução
No mundo tecnológico moderno aumentou-se a necessidade de uma educação formal que habilite o indivíduo ao mercado de trabalho e lhe permita usufruir melhor das oportunidades de lazer e saúde disponíveis na sociedade. Essa demanda de educação formal, por sua vez, pressiona a sociedade para que melhore seus serviços de ensino, quer público ou privado, para prover uma educação que não só beneficie o indivíduo, mas que também garanta o desenvolvimento tecnológico futuro dessa mesma sociedade. A demanda pela educação inclui grupos de indivíduos com necessidades especiais decorrentes de suas condições físicas e biológicas, que acarretam distúrbios ou deficiências de aprendizagem.
Cerca de dez por cento da população em idade escolar experimenta uma disfunção cerebral que resulta em um distúrbio ou deficiência de aprendizagem (Aicardi, 1998; Capute e Accardo, 1996; Coffey and Brumback, 1998; Spreen et al, 1995). Entre 0.56 a 2.3 por 1000 crianças apresentam perda sensível da audição que compromete o desenvolvimento da linguagem falada (Panoscha, 1996). Embora essas crianças tenham sido discriminadas no passado, hoje se procura incluí-las no processo educacional. Para isso torna-se necessária uma melhor compreensão de suas capacidades e de como utilizá-las no processo de ensino.
O uso dos novos conhecimentos disponibilizados pelas neurociências na modelagem dos processos formais educativos tem resultado em novas propostas de trabalho nas áreas da educação infantil e ensino fundamental (Butterworth, 1999; Cardoso-Martins, 1996; Harley, 1995; Pinheiro, 1994; Siegler, 1996; Rocha, 1999, 2000). Essas novas abordagens têm contribuído para uma melhoria da qualidade de ensino e um aumento de sua efetividade. Assim, para que novas propostas de trabalhos sejam criadas é fundamental primeiro compreender o processamento neural da criança com deficiência, para com isso saber quais são os recursos a serem utilizados para facilitar seu aprendizado.
Todas as sociedades humanas utilizam gestos motores para complementarem suas informações sonoras veiculadas por meio da fala. Muitas vezes nos parece impossível falar sem movermos nossas mãos, nossos olhos e nossos braços (Armstrong et al. 1995). Para os falantes esses gestos apenas ajudam a ilustrar um objeto, indicar uma direção, mostrar um estado emocional, mas para indivíduos privados da percepção do estímulo sonoro, eles passam a ser a única expressão capaz de ser compreendida. Esses indivíduos aprimoram tanto seus gestos que se tornam capazes de se comunicarem exclusivamente por meio de uma linguagem de sinais.
As línguas de sinais surgem de maneira espontânea, pela utilização de gestos e por mímicas realizadas por um grupo de indivíduos surdos (Capovila et al, 1998). Tal grupo convenciona e desenvolve certos padrões gestuais próprios para cada objeto, ação, estado psíquico e emocional. O aperfeiçoamento de tais padrões alcança a complexidade existente em qualquer língua falada, contendo todos os níveis lingüísticos: fonológico, morfológico, sintático, semântico e pragmático.
Para a língua brasileira de sinais (Libras), como para outras linguagens por sinais, a fonologia delimita as unidades mínimas distintivas, consideradas como fonemas, a partir da decomposição dos movimentos das mãos, dos braços e das expressões faciais. Brito (1995) divide os fonemas manuais em duas categorias, baseando-se nos seguintes parâmetros: primários, que definem a configuração das mãos, sua posição em relação ao corpo (ponto de articulação) e o movimento exercido por elas; secundários, que se referem à região de contato das mãos, à orientação das mesmas e à disposição das palmas (para baixo ou para cima). As expressões faciais e os movimentos do corpo não chegam a distinguir significados distintos, mas funcionam como modificadores.
A morfologia da língua de sinais também pode ser segmentada em fonemas. Algumas palavras são formadas apenas por uma determinada configuração de mão apresentada em qualquer posição do espaço, porém a maioria das palavras nas línguas de sinais é formada por um conjunto visual que pode envolver todos os parâmetros distintivos. Na sintaxe, a Libras se faz compreender pela ordem temporal dos constituintes durante a sinalização. A ordem padrão é sujeito, objeto e verbo, mas como nas línguas faladas tal ordem pode ser trocada de acordo com a topicalização desejada, contanto que não se crie ambigüidade.
Sabe-se que no processamento e produção da fala os falantes usam áreas específicas do lóbulo temporal esquerdo e de áreas frontais esquerdas. No processamento cerebral de uma língua falada o estímulo sensorial sonoro é codificado primeiramente na área auditiva verbal, onde as informações sensoriais referentes a cada palavra fazem certos neurônios dessa área se associarem aos neurônios da área de Broca e certos neurônios à área de Wernicke (van Berkum et al. 1999; Cohen et al. 2000; Federmeier et al, 2000; Murta et al, 1999; Perani et al. 1999; Ni et al, 2000; Rocha 1999; Rocha 2000; Tarkiainen et al. 1999). As palavras que denominam uma ação (verbo) são codificadas por neurônios da área de Broca, que representa os movimentos dos verbos. As palavras que definem um nome (substantivo e adjetivo) são codificadas pelos neurônios da área de Wernicke, que por sua vez se associam com os neurônios ou das áreas visuais para composição de um objeto, ou da área límbica para identificação de um sentimento, ou das áreas de olfação e gustação para recriação das sensações olfativas e palatais. Isto é, por meio de Wernicke ativam-se neurônios que estão associados às características semânticas das palavras (Damasio et al. 1996).
Pode-se criar a hipótese de que os verbos são inicialmente identificados em Broca, sendo ativadas a partir dele áreas parieto-temporais para o recrutamento das palavras adequadas referentes à semântica e à sintaxe de cada verbo (Rocha et al, 2001). Rizzolatti and Arbib (1998) propuseram que Broca é uma área análoga a uma área frontal do cérebro dos macacos, onde os autores encontraram "neurônios espelhos", ou seja, células que são ativadas quando os macacos estão executando uma ação ou quando estão apenas observando outro indivíduo realizar uma ação semelhante. A área de Broca possui, portanto, neurônios que respondem tanto à ação motora em si quanto à informação sensorial associada à mesma ação, sendo ela auditiva ou visual. No primeiro caso um neurônio representa uma ação no sistema motor e será chamado, aqui, de neurônio de representação de ações; no segundo caso representa a fonação de uma palavra e será chamado, aqui, de neurônio de representação verbal. A proximidade desses dois neurônios na área de Broca facilita as conexões sinápticas entre eles, e consequentemente cria a condição para definir a sintaxe e semântica (neurônio de representação da ação) do verbo (neurônio de representação verbal). Um modelo semelhante pode ser assumido para a área de Wernicke, onde neurônios que possuem conexões com as áreas de processamento das outras sensações físicas: visão, tato, olfato, e com as áreas de memória, do sistema límbico, etc., possibilitariam o acesso à semântica das palavras representadas verbalmente em neurônios do córtex auditivo vizinho ou mesmo em neurônios da própria área de Wernicke.
Na figura 1, por exemplo, os neurônios auditivos temporais que reconhecem a forma auditiva da palavra comer, ativam neurônios na área de Broca (come na Fig 1) correspondentes à sua fonação e à ação representada por esse verbo, e o reconhecimento das formas sonoras das palavras leão e carne ativa os neurônios da área de Wernicke que dão acesso às áreas cerebrais que reconhecem esses elementos e definem a semântica dessas palavras. O neurônio de representação das ações na área de Broca estabelece relações sinápticas com neurônios de representação dos nomes em Wernicke, que representem elementos que podem estar envolvidos com a ação que o verbo descreve. Dessa maneira o reconhecimento do verbo comer na área de Broca, favorece a ativação dos neurônios referentes às palavras leão e comer, o que confirma que essas palavras obedecem a sintaxe do verbo comer e fazem parte do campo semântico da ação desse verbo.
(LÍNGUA BRASILEIRA DE SINAIS)
UM ESTUDO ELETROENCEFALOGRAFICO DE SUA FUNCIONALIDADE CEREBRAL
Fábio Theoto Rocha
Gerente da EINA - Estudos em Inteligência Natural e Artificial Ltda
Introdução
No mundo tecnológico moderno aumentou-se a necessidade de uma educação formal que habilite o indivíduo ao mercado de trabalho e lhe permita usufruir melhor das oportunidades de lazer e saúde disponíveis na sociedade. Essa demanda de educação formal, por sua vez, pressiona a sociedade para que melhore seus serviços de ensino, quer público ou privado, para prover uma educação que não só beneficie o indivíduo, mas que também garanta o desenvolvimento tecnológico futuro dessa mesma sociedade. A demanda pela educação inclui grupos de indivíduos com necessidades especiais decorrentes de suas condições físicas e biológicas, que acarretam distúrbios ou deficiências de aprendizagem.
Cerca de dez por cento da população em idade escolar experimenta uma disfunção cerebral que resulta em um distúrbio ou deficiência de aprendizagem (Aicardi, 1998; Capute e Accardo, 1996; Coffey and Brumback, 1998; Spreen et al, 1995). Entre 0.56 a 2.3 por 1000 crianças apresentam perda sensível da audição que compromete o desenvolvimento da linguagem falada (Panoscha, 1996). Embora essas crianças tenham sido discriminadas no passado, hoje se procura incluí-las no processo educacional. Para isso torna-se necessária uma melhor compreensão de suas capacidades e de como utilizá-las no processo de ensino.
O uso dos novos conhecimentos disponibilizados pelas neurociências na modelagem dos processos formais educativos tem resultado em novas propostas de trabalho nas áreas da educação infantil e ensino fundamental (Butterworth, 1999; Cardoso-Martins, 1996; Harley, 1995; Pinheiro, 1994; Siegler, 1996; Rocha, 1999, 2000). Essas novas abordagens têm contribuído para uma melhoria da qualidade de ensino e um aumento de sua efetividade. Assim, para que novas propostas de trabalhos sejam criadas é fundamental primeiro compreender o processamento neural da criança com deficiência, para com isso saber quais são os recursos a serem utilizados para facilitar seu aprendizado.
Todas as sociedades humanas utilizam gestos motores para complementarem suas informações sonoras veiculadas por meio da fala. Muitas vezes nos parece impossível falar sem movermos nossas mãos, nossos olhos e nossos braços (Armstrong et al. 1995). Para os falantes esses gestos apenas ajudam a ilustrar um objeto, indicar uma direção, mostrar um estado emocional, mas para indivíduos privados da percepção do estímulo sonoro, eles passam a ser a única expressão capaz de ser compreendida. Esses indivíduos aprimoram tanto seus gestos que se tornam capazes de se comunicarem exclusivamente por meio de uma linguagem de sinais.
As línguas de sinais surgem de maneira espontânea, pela utilização de gestos e por mímicas realizadas por um grupo de indivíduos surdos (Capovila et al, 1998). Tal grupo convenciona e desenvolve certos padrões gestuais próprios para cada objeto, ação, estado psíquico e emocional. O aperfeiçoamento de tais padrões alcança a complexidade existente em qualquer língua falada, contendo todos os níveis lingüísticos: fonológico, morfológico, sintático, semântico e pragmático.
Para a língua brasileira de sinais (Libras), como para outras linguagens por sinais, a fonologia delimita as unidades mínimas distintivas, consideradas como fonemas, a partir da decomposição dos movimentos das mãos, dos braços e das expressões faciais. Brito (1995) divide os fonemas manuais em duas categorias, baseando-se nos seguintes parâmetros: primários, que definem a configuração das mãos, sua posição em relação ao corpo (ponto de articulação) e o movimento exercido por elas; secundários, que se referem à região de contato das mãos, à orientação das mesmas e à disposição das palmas (para baixo ou para cima). As expressões faciais e os movimentos do corpo não chegam a distinguir significados distintos, mas funcionam como modificadores.
A morfologia da língua de sinais também pode ser segmentada em fonemas. Algumas palavras são formadas apenas por uma determinada configuração de mão apresentada em qualquer posição do espaço, porém a maioria das palavras nas línguas de sinais é formada por um conjunto visual que pode envolver todos os parâmetros distintivos. Na sintaxe, a Libras se faz compreender pela ordem temporal dos constituintes durante a sinalização. A ordem padrão é sujeito, objeto e verbo, mas como nas línguas faladas tal ordem pode ser trocada de acordo com a topicalização desejada, contanto que não se crie ambigüidade.
Sabe-se que no processamento e produção da fala os falantes usam áreas específicas do lóbulo temporal esquerdo e de áreas frontais esquerdas. No processamento cerebral de uma língua falada o estímulo sensorial sonoro é codificado primeiramente na área auditiva verbal, onde as informações sensoriais referentes a cada palavra fazem certos neurônios dessa área se associarem aos neurônios da área de Broca e certos neurônios à área de Wernicke (van Berkum et al. 1999; Cohen et al. 2000; Federmeier et al, 2000; Murta et al, 1999; Perani et al. 1999; Ni et al, 2000; Rocha 1999; Rocha 2000; Tarkiainen et al. 1999). As palavras que denominam uma ação (verbo) são codificadas por neurônios da área de Broca, que representa os movimentos dos verbos. As palavras que definem um nome (substantivo e adjetivo) são codificadas pelos neurônios da área de Wernicke, que por sua vez se associam com os neurônios ou das áreas visuais para composição de um objeto, ou da área límbica para identificação de um sentimento, ou das áreas de olfação e gustação para recriação das sensações olfativas e palatais. Isto é, por meio de Wernicke ativam-se neurônios que estão associados às características semânticas das palavras (Damasio et al. 1996).
Pode-se criar a hipótese de que os verbos são inicialmente identificados em Broca, sendo ativadas a partir dele áreas parieto-temporais para o recrutamento das palavras adequadas referentes à semântica e à sintaxe de cada verbo (Rocha et al, 2001). Rizzolatti and Arbib (1998) propuseram que Broca é uma área análoga a uma área frontal do cérebro dos macacos, onde os autores encontraram "neurônios espelhos", ou seja, células que são ativadas quando os macacos estão executando uma ação ou quando estão apenas observando outro indivíduo realizar uma ação semelhante. A área de Broca possui, portanto, neurônios que respondem tanto à ação motora em si quanto à informação sensorial associada à mesma ação, sendo ela auditiva ou visual. No primeiro caso um neurônio representa uma ação no sistema motor e será chamado, aqui, de neurônio de representação de ações; no segundo caso representa a fonação de uma palavra e será chamado, aqui, de neurônio de representação verbal. A proximidade desses dois neurônios na área de Broca facilita as conexões sinápticas entre eles, e consequentemente cria a condição para definir a sintaxe e semântica (neurônio de representação da ação) do verbo (neurônio de representação verbal). Um modelo semelhante pode ser assumido para a área de Wernicke, onde neurônios que possuem conexões com as áreas de processamento das outras sensações físicas: visão, tato, olfato, e com as áreas de memória, do sistema límbico, etc., possibilitariam o acesso à semântica das palavras representadas verbalmente em neurônios do córtex auditivo vizinho ou mesmo em neurônios da própria área de Wernicke.
Na figura 1, por exemplo, os neurônios auditivos temporais que reconhecem a forma auditiva da palavra comer, ativam neurônios na área de Broca (come na Fig 1) correspondentes à sua fonação e à ação representada por esse verbo, e o reconhecimento das formas sonoras das palavras leão e carne ativa os neurônios da área de Wernicke que dão acesso às áreas cerebrais que reconhecem esses elementos e definem a semântica dessas palavras. O neurônio de representação das ações na área de Broca estabelece relações sinápticas com neurônios de representação dos nomes em Wernicke, que representem elementos que podem estar envolvidos com a ação que o verbo descreve. Dessa maneira o reconhecimento do verbo comer na área de Broca, favorece a ativação dos neurônios referentes às palavras leão e comer, o que confirma que essas palavras obedecem a sintaxe do verbo comer e fazem parte do campo semântico da ação desse verbo.

Além de Broca e Wernicke há uma região específica para codificação das palavras de função (Berkum et al, 1999; Brown et al. 1999). A posição de tal área ainda não está bem definida, mas supõe-se que ela se encontra próxima à área de Broca. Essa área é fundamental para a análise da relação das palavras em uma frase, pois, fora a ordem das palavras, é por meio das conjunções e preposições que podemos definir a função delas na frase. No lóbulo frontal a ordem das palavras e as informações veiculadas pelas palavras de função fazem com que as palavras da frase façam sentido, uma em relação à outra e permitam a compreensão do enunciado como um todo.
A decodificação verbal deve acessar memórias distintas para extrair significado das informações recebidas visual ou verbalmente (Smith et al, 1998), algumas vezes de forma ordenada no tempo; algumas vezes de acordo com relações espaciais definidas; algumas vezes dependendo da familiaridade, etc.(Curtis et al, 2000; Dafner et al, 2000; Gabrieli, et al, 1998; Elias et al, 1999; Hopfinger et al, 2000; Marshetz et al, 2000; Smith et al, 1998; St George et al, 1999; Ungerleider et al, 1998). A decodificação verbal é, portanto, uma tarefa complicada que deve ser resolvida por um grande número de neurônios distribuídos por quase todo o cérebro, neurônios esses especializados em tratar os diferentes aspectos do discurso. O recrutamento da memória, o controle da atenção, o processamento visual e a imaginação mental envolvida nesse processo, são basicamente controlados pelo córtex frontal, mas também envolve neurônios distribuídos por outras áreas: parietal, temporal e occipital, além do hipocampo e áreas adjacentes (Brewer et al, 1998; Henson et al, 1999; Hopfinger et al, 2000; Kosslyn et al, 1999; McDermott et al, 2000; Rocha et al, 2001).
A literatura tem mostrado até o instante que os circuitos neurais para a linguagem de sinais funcionam de maneira, senão idêntica, ao menos semelhante no processamento cerebral da língua oral. Porém, a diferença clara e fundamental é a natureza do estímulo sensorial. As áreas lingüísticas não sendo mais ativadas a partir da área auditiva verbal serão ativadas a partir de áreas de processamento visual (Nishimura et al, 1999; Söderfeldt et al, 1997; Emmorey et al, 2001). As áreas visuais primárias se associam a área de representação de movimento das mãos (que se encontra na região parietal esquerda). Essa área passa a fornecer a informação necessária para Wernicke e Broca codificarem a ação dos verbos e a semântica dos nomes. Outras áreas visuais serão recrutadas para a identificação da sua relação em relação ao corpo, ao seu formato e sua direção. (Ishai et al, 2000; Kanwisher et al, 1997; Maunsell et al, 1995; Nakamura et al, 2000; O’Craven et al, 2000; Riesenhuber et al, 1999; Young, 1995; Emmorey et al, 2001). Essas áreas se acham distribuídas no córtex temporal bilateralmente.
Outra diferença marcante nas línguas de sinais é o uso do espaço como referência anafórica. Assim, projeta-se um objeto em uma determinada posição do espaço e quando se quer retomar esse objeto na frase aponta-se para a posição em que ele havia sido transportado (Bellugi et al, 1996). Tal recurso cria a necessidade de outras áreas cerebrais, referentes ao processamento visual espacial, se envolverem no processamento lingüístico. A incapacidade de controlar tal processo acarreta aos sinalizantes um tipo de afasia onde o indivíduo perde a capacidade de manter um discurso coerente. Segundo Hickok et al. (1999), um indivíduo com lesões nas áreas mais anteriores do hemisfério direito era incapaz de utilizar a referência espacial e manter um discurso coerente, contudo ele ainda mantinha operante todas as outras funções lingüísticas. A referência espacial depende praticamente do processamento neural do campo visual, relacionando os objetos uns em relação aos outros de acordo com sua posição no espaço. Tal processamento ocorre justamente nas áreas mais anteriores do hemisfério direito, por isso a afazia de discurso nos pacientes com lesões de hemisfério direito.
A organização estrutural das redes neurais referentes a cada processamento cognitivo, apesar de estar predeterminada pela genética, também depende dos estímulos externos recebidos, principalmente, durante a infância (Neville et al., 1997). Analisando a atividade cerebral de indivíduos surdos e ouvintes que haviam aprendido a língua de sinais na infância, Neville observa ativações semelhantes àquelas ocorridas no processamento das línguas faladas. No entanto, quando analisa a atividade cerebral de indivíduos que aprenderam a língua de sinais tardiamente, nota ativações diferentes, pois tal língua não foi incorporada pelo mesmo sistema lingüístico da criança surda, o que acarreta um déficit na performance daqueles indivíduos. A surdez congênita induz modificações na estruturação cerebral, pois elimina a competição sináptica entre informações visuais e verbais em várias regiões do cérebro. Tal surdez acarreta uma diminuição das terminações nervosas sonoras, e um conseqüente aumento das terminações nervosas visuais. Essa recompensa faz com que indivíduos surdos adquiram maior capacidade computacional do campo visual periférico com a utilização de áreas temporais. Essas áreas normalmente estão relacionadas ao processamento sonoro, mas nos surdos, por falta do estímulo físico, passam a processar informações visuais refletidas nas margens periférica da retina (Neville, 1996).
Objetivos.
O objetivo básico do presente trabalho é procurar compreender o processo neurofisiológico empregado pelos surdos durante o processamento da Libras, analisando a atividade elétrica cerebral através da tecnologia de Mapeamento Cognitivo Cerebral (MCC), desenvolvido por Rocha et al (2001). Pretende-se também estudar as possíveis diferenças de ativação cerebral no processamento da Libras entre surdos e ouvintes sinalizantes. Esse estudo visa orientar o processo didático dos surdos, mostrando os caminhos mais fáceis e produtivos para o cérebro de uma criança surda aprender a se comunicar com outros surdos e com ouvintes não sinalizantes através da escrita.
Com isso pretende-se contribuir para um melhor conhecimento da Libras e para o desenvolvimento de estratégias de ensino mais adequadas aos surdos, aumentando a consciência da rede educacional brasileira para a existência de uma nova língua atuante no Brasil e carente de um apoio pedagógico apropriado.
Metodologia.
Uma história folclórica do Currupira e um conjunto de charadas sobre animais foram utilizados para investigar a atividade elétrica cerebral associada à compreensão da Libras. A sinalização da história e das charadas foi gravada em vídeo, por uma sinalizadora da Libras (Fig. 2). A história foi dividida em quatro trechos, correspondente à descrição do personagem Currupira, do seu comportamento e de sua função na floresta. Cada um dos trechos da história e das charadas foi ilustrado por um desenhista profissional. O vídeo e as ilustrações foram utilizados para montagem de videojogos do sistema ENSCER (Rocha, 2000), chamados aqui de História e Charadas (Fig. 3). Esses jogos com as respectivas ilustrações e sinalizações encontram-se no CD Rom anexado.
Os vídeos correspondentes a cada trecho da história ou da charada eram apresentados em um monitor A, enquanto que as figuras correspondentes eram apresentadas em outro monitor B (Fig. 4). Ambos os vídeos foram ligados ao mesmo computador que controlava os jogos. Após a visualização de cada vídeo os voluntários deviam escolher a cena correspondente ao trecho da história descrito no vídeo, ou ao animal descrito na sinalização da charada.
O estudo da atividade elétrica cerebral associada à compreensão da LIBRAS utilizou voluntários destros: 10 crianças surdas congênitas sinalizadoras, idade média de 12 anos, e 5 adultos em processo de aquisição da Libras, idade média de 28 anos. Com nenhuma história de distúrbios neurológicos.
A decodificação verbal deve acessar memórias distintas para extrair significado das informações recebidas visual ou verbalmente (Smith et al, 1998), algumas vezes de forma ordenada no tempo; algumas vezes de acordo com relações espaciais definidas; algumas vezes dependendo da familiaridade, etc.(Curtis et al, 2000; Dafner et al, 2000; Gabrieli, et al, 1998; Elias et al, 1999; Hopfinger et al, 2000; Marshetz et al, 2000; Smith et al, 1998; St George et al, 1999; Ungerleider et al, 1998). A decodificação verbal é, portanto, uma tarefa complicada que deve ser resolvida por um grande número de neurônios distribuídos por quase todo o cérebro, neurônios esses especializados em tratar os diferentes aspectos do discurso. O recrutamento da memória, o controle da atenção, o processamento visual e a imaginação mental envolvida nesse processo, são basicamente controlados pelo córtex frontal, mas também envolve neurônios distribuídos por outras áreas: parietal, temporal e occipital, além do hipocampo e áreas adjacentes (Brewer et al, 1998; Henson et al, 1999; Hopfinger et al, 2000; Kosslyn et al, 1999; McDermott et al, 2000; Rocha et al, 2001).
A literatura tem mostrado até o instante que os circuitos neurais para a linguagem de sinais funcionam de maneira, senão idêntica, ao menos semelhante no processamento cerebral da língua oral. Porém, a diferença clara e fundamental é a natureza do estímulo sensorial. As áreas lingüísticas não sendo mais ativadas a partir da área auditiva verbal serão ativadas a partir de áreas de processamento visual (Nishimura et al, 1999; Söderfeldt et al, 1997; Emmorey et al, 2001). As áreas visuais primárias se associam a área de representação de movimento das mãos (que se encontra na região parietal esquerda). Essa área passa a fornecer a informação necessária para Wernicke e Broca codificarem a ação dos verbos e a semântica dos nomes. Outras áreas visuais serão recrutadas para a identificação da sua relação em relação ao corpo, ao seu formato e sua direção. (Ishai et al, 2000; Kanwisher et al, 1997; Maunsell et al, 1995; Nakamura et al, 2000; O’Craven et al, 2000; Riesenhuber et al, 1999; Young, 1995; Emmorey et al, 2001). Essas áreas se acham distribuídas no córtex temporal bilateralmente.
Outra diferença marcante nas línguas de sinais é o uso do espaço como referência anafórica. Assim, projeta-se um objeto em uma determinada posição do espaço e quando se quer retomar esse objeto na frase aponta-se para a posição em que ele havia sido transportado (Bellugi et al, 1996). Tal recurso cria a necessidade de outras áreas cerebrais, referentes ao processamento visual espacial, se envolverem no processamento lingüístico. A incapacidade de controlar tal processo acarreta aos sinalizantes um tipo de afasia onde o indivíduo perde a capacidade de manter um discurso coerente. Segundo Hickok et al. (1999), um indivíduo com lesões nas áreas mais anteriores do hemisfério direito era incapaz de utilizar a referência espacial e manter um discurso coerente, contudo ele ainda mantinha operante todas as outras funções lingüísticas. A referência espacial depende praticamente do processamento neural do campo visual, relacionando os objetos uns em relação aos outros de acordo com sua posição no espaço. Tal processamento ocorre justamente nas áreas mais anteriores do hemisfério direito, por isso a afazia de discurso nos pacientes com lesões de hemisfério direito.
A organização estrutural das redes neurais referentes a cada processamento cognitivo, apesar de estar predeterminada pela genética, também depende dos estímulos externos recebidos, principalmente, durante a infância (Neville et al., 1997). Analisando a atividade cerebral de indivíduos surdos e ouvintes que haviam aprendido a língua de sinais na infância, Neville observa ativações semelhantes àquelas ocorridas no processamento das línguas faladas. No entanto, quando analisa a atividade cerebral de indivíduos que aprenderam a língua de sinais tardiamente, nota ativações diferentes, pois tal língua não foi incorporada pelo mesmo sistema lingüístico da criança surda, o que acarreta um déficit na performance daqueles indivíduos. A surdez congênita induz modificações na estruturação cerebral, pois elimina a competição sináptica entre informações visuais e verbais em várias regiões do cérebro. Tal surdez acarreta uma diminuição das terminações nervosas sonoras, e um conseqüente aumento das terminações nervosas visuais. Essa recompensa faz com que indivíduos surdos adquiram maior capacidade computacional do campo visual periférico com a utilização de áreas temporais. Essas áreas normalmente estão relacionadas ao processamento sonoro, mas nos surdos, por falta do estímulo físico, passam a processar informações visuais refletidas nas margens periférica da retina (Neville, 1996).
Objetivos.
O objetivo básico do presente trabalho é procurar compreender o processo neurofisiológico empregado pelos surdos durante o processamento da Libras, analisando a atividade elétrica cerebral através da tecnologia de Mapeamento Cognitivo Cerebral (MCC), desenvolvido por Rocha et al (2001). Pretende-se também estudar as possíveis diferenças de ativação cerebral no processamento da Libras entre surdos e ouvintes sinalizantes. Esse estudo visa orientar o processo didático dos surdos, mostrando os caminhos mais fáceis e produtivos para o cérebro de uma criança surda aprender a se comunicar com outros surdos e com ouvintes não sinalizantes através da escrita.
Com isso pretende-se contribuir para um melhor conhecimento da Libras e para o desenvolvimento de estratégias de ensino mais adequadas aos surdos, aumentando a consciência da rede educacional brasileira para a existência de uma nova língua atuante no Brasil e carente de um apoio pedagógico apropriado.
Metodologia.
Uma história folclórica do Currupira e um conjunto de charadas sobre animais foram utilizados para investigar a atividade elétrica cerebral associada à compreensão da Libras. A sinalização da história e das charadas foi gravada em vídeo, por uma sinalizadora da Libras (Fig. 2). A história foi dividida em quatro trechos, correspondente à descrição do personagem Currupira, do seu comportamento e de sua função na floresta. Cada um dos trechos da história e das charadas foi ilustrado por um desenhista profissional. O vídeo e as ilustrações foram utilizados para montagem de videojogos do sistema ENSCER (Rocha, 2000), chamados aqui de História e Charadas (Fig. 3). Esses jogos com as respectivas ilustrações e sinalizações encontram-se no CD Rom anexado.
Os vídeos correspondentes a cada trecho da história ou da charada eram apresentados em um monitor A, enquanto que as figuras correspondentes eram apresentadas em outro monitor B (Fig. 4). Ambos os vídeos foram ligados ao mesmo computador que controlava os jogos. Após a visualização de cada vídeo os voluntários deviam escolher a cena correspondente ao trecho da história descrito no vídeo, ou ao animal descrito na sinalização da charada.
O estudo da atividade elétrica cerebral associada à compreensão da LIBRAS utilizou voluntários destros: 10 crianças surdas congênitas sinalizadoras, idade média de 12 anos, e 5 adultos em processo de aquisição da Libras, idade média de 28 anos. Com nenhuma história de distúrbios neurológicos.

Fig. 2 Uma amostra do vídeo com a sinalização da LIBRAS

A

B
Fig. 3 Figuras utilizadas no videojogo Histórias (A) e Charadas (B)
Utilizou-se dois computadores para realizar o estudo (Figura 4). Um deles fez a aquisição e o registro dos sinais eletroencefalográficos e o outro apresentou os videojogos. Os dois computadores operaram em rede, de modo a permitir a sincronização da aquisição do EEG com os eventos associados às distintas fases dos vídeos jogos. Para colocação dos eletrodos utilizou-se o sistema 10/20, como ilustrado na figura 4, e a aquisição do EEG é feita a uma freqüência de 256 Hz e com uma precisão de 10 bits.
Os 20 eletrodos são fixados com ajuda de pasta condutora e distribuídos pelo crânio, 7 na região frontal (FP1; FP2, F7; F3; FZ; F4; F8), 3 na região central (C3; CZ; C4), 3 parietais (P3; PZ; P4), 4 temporais (T3; T5; T4; T6) e 3 occipitais (O1; OZ; O2). São utilizados também 3 eletrodos ( 1 para terra e duas referências), localizados 1 na fronte e 2 nos lóbulos da orelha, conforme ilustrado na figura 4.
Os 20 eletrodos são fixados com ajuda de pasta condutora e distribuídos pelo crânio, 7 na região frontal (FP1; FP2, F7; F3; FZ; F4; F8), 3 na região central (C3; CZ; C4), 3 parietais (P3; PZ; P4), 4 temporais (T3; T5; T4; T6) e 3 occipitais (O1; OZ; O2). São utilizados também 3 eletrodos ( 1 para terra e duas referências), localizados 1 na fronte e 2 nos lóbulos da orelha, conforme ilustrado na figura 4.
A atividade elétrica cerebral associada a uma tarefa cognitiva registrada pelo EEG, pode ser estudada através da técnica da Atividade Relacionada a Eventos (ARE) (veja, por exemplo, Deary et al, 1997, Hillyard et al, 1998, Rocha, 1990). Nessa técnica, determina-se quais eventos associados à determinada tarefa cognitiva serão estudados. A atividade elétrica cerebral durante um período pré-definido é registrada e sua média calculada para todas as ocorrências desses eventos durante o teste a que o grupo de indivíduos é submetido. Calcula-se, portanto, a AREe,k associada ao evento e do videojogo k (Fig. 4) para todos os m voluntários de cada grupo experimental. Esse cálculo foi realizado para um período de 2000 ms, para os seguintes eventos:
Apresentação da história: 0, 2 e 4 segundos após o início da apresentação, a conectividade média calculada para todo o período da apresentação, dois segundos antes e dois segundos depois do término da apresentação.
Charadas: 0, 2 e 4 segundos após o início da apresentação, a conectividade média calculada para todo o período da apresentação, dois segundos antes e dois segundos depois do término da apresentação, e no momento da escolha da decisão correta.
Após a obtenção da AREe,k para o evento e do videojogo k, calcula-se a correlação ri,j entre a atividade promediada para cada um dos eletrodos ( i ) com os outros 19 eletrodos ( j ), e assim sucessivamente. Utiliza-se esse coeficiente de correlação linear ri,j para a geração do correspondente Mapa Cognitivo Cerebral (MCCe,k) de acordo com os seguintes cálculos (Rocha et al, 2001b):
a) a entropia de associação h(ai,j) entre a atividade elétrica promediada para os eletrodos i,j é obtida como:
h(ai,j) = - ri,j log2 ri,j - ( 1 - ri,j ) log2 ( 1 - ri,j )
b) a entropia média de associação h(mi) entre o eletrodo i os outros 19 eletrodos j é obtida como:
h(mi) = - ri log2 ri - ( 1 - ri ) log2 ( 1 - ri )
onde:
20
ri = 1/20 S ri,j
j=1
c) a entropia de comunicação h(ci) utilizada pelos neurônios registrados pelo eletrodo i é obtida como:
20
h(ci) = S h(mi) - h(ai,j)
j=1
O MCCe,k para o evento e do videojogo k é um gráfico em que os valores de h(ci) para cada eletrodo i é normalizado e codificado em cores conforme escala previamente definida (Fig. 4), e plotado de acordo com a distribuição espacial dos eletrodos de registro.
Resumindo o procedimento para realizar tal estudo:
registra-se o EEG durante a realização dos videojogos,
utilizam-se dois computadores ligados em rede: um para o registro do EEG e outro para a execução das atividades
os dois computadores são sincronizados, de modo que o EEG registrado é referenciado aos diversos eventos para cada um dos vídeos jogos;
selecionam-se os trechos do EEG associados a cada um dos eventos de cada jogo.
calcula-se para cada evento da atividade e para cada grupo experimento, a média da atividade elétrica relacionada a esse evento (ARE);
obtém-se a correlação linear da atividade promediada para cada derivação dos AREs em relação à atividade promediada de todas as outras 19 derivações;
calcula-se então a capacidade computacional (h(c)), ou seja, a entropia alocada em cada área ou derivação do EEG para a execução do vídeo jogo, e finalmente
constrói-se os mapas cognitivos cerebrais ( MCCs ) mostrando a capacidade computacional (h(c)) alocada em cada derivação para a execução de um evento da tarefa estudada.
Usou-se, também, análise fatorial para estudar as possíveis correlações de variança entre os valores de h(c) calculados para as distintas derivações do EEG. Dois fatores foram extraidos usando a técnica de Componente Principais, e esses fatores foram rotacionados usando-se o método Varimax Normalizado. Se esses dois fatores explicassem mais de 50% da variabilidade total da h(c), então a análise era considerada válida, e mapas cerebrais eram construídos (chamados aqui de MFs – figs.) codificando em cores os valores do coeficiente de correlação obtido para cada uma das derivações do EEG.
Resultados.
O início da apresentação de cada trecho da história do Currupira (fig. 5) aos surdos recrutou neurônios das áreas centrais (CZ, C3, C4) e frontais (F3, FP2, F4) de ambos os hemisférios, e da área parietal medial (PZ). Após dois segundos de apresentação da história observa-se o envolvimento predominante de neurônios na região frontal e parietal do hemisfério esquerdo (F3, P3), na região frontal e central medial (FZ, CZ), e na região temporal direita (T6). Após quatro segundos o recrutamento envolve áreas bilaterais frontais (F7, F8), áreas mediais da região parietal e central (PZ, CZ), e ainda a área frontal direita (F4). A conectividade média calculada para todo o período de sinalização, mostrou envolvimento de ampla área frontal (F3, F4, FZ, F8), parietal (PZ, P3, P4), e central (C3, C4, CZ), além de OZ. Dois segundos antes do termino da sinalização as mesmas áreas estão ativadas com exceção de FZ, P3 e F8, que deixam de serem recrutadas, e de T6 que passa a ser ativada. Dois segundos após a sinalização as únicas áreas não ativadas são O2, T5, T6, T4, T3, P4 e FP1.
No jogo de charada (fig. 6) foi adicionado mais um evento: A – o momento de escolha da resposta correta. O início da apresentação da informação sinalizada recrutou células das áreas centrais do cérebro (CZ, C3, C4), frontais (F3), com um predomínio do hemisfério direito (F4, F8, FP2), e parietais (PZ). Após dois segundos de sinalização, observou-se uma maior participação de neurônios de ambos os hemisférios nas áreas frontais (F3, F4, F8, F7,FP2), além de neurônios temporais e occipitais direitos (T4, T6, O2). Após quatro segundos a ativação bilateral permanece nas áreas frontais, nas regiões dos eletrodos F3 e F4, e passa também a recrutar bilateralmente as áreas occipitais (O1, O2) e temporais (T5, T6), no hemisfério esquerdo há ainda a ativação de neurônios temporais e centrais (T3, C3), na região medial ocorre ativação de CZ.
Apresentação da história: 0, 2 e 4 segundos após o início da apresentação, a conectividade média calculada para todo o período da apresentação, dois segundos antes e dois segundos depois do término da apresentação.
Charadas: 0, 2 e 4 segundos após o início da apresentação, a conectividade média calculada para todo o período da apresentação, dois segundos antes e dois segundos depois do término da apresentação, e no momento da escolha da decisão correta.
Após a obtenção da AREe,k para o evento e do videojogo k, calcula-se a correlação ri,j entre a atividade promediada para cada um dos eletrodos ( i ) com os outros 19 eletrodos ( j ), e assim sucessivamente. Utiliza-se esse coeficiente de correlação linear ri,j para a geração do correspondente Mapa Cognitivo Cerebral (MCCe,k) de acordo com os seguintes cálculos (Rocha et al, 2001b):
a) a entropia de associação h(ai,j) entre a atividade elétrica promediada para os eletrodos i,j é obtida como:
h(ai,j) = - ri,j log2 ri,j - ( 1 - ri,j ) log2 ( 1 - ri,j )
b) a entropia média de associação h(mi) entre o eletrodo i os outros 19 eletrodos j é obtida como:
h(mi) = - ri log2 ri - ( 1 - ri ) log2 ( 1 - ri )
onde:
20
ri = 1/20 S ri,j
j=1
c) a entropia de comunicação h(ci) utilizada pelos neurônios registrados pelo eletrodo i é obtida como:
20
h(ci) = S h(mi) - h(ai,j)
j=1
O MCCe,k para o evento e do videojogo k é um gráfico em que os valores de h(ci) para cada eletrodo i é normalizado e codificado em cores conforme escala previamente definida (Fig. 4), e plotado de acordo com a distribuição espacial dos eletrodos de registro.
Resumindo o procedimento para realizar tal estudo:
registra-se o EEG durante a realização dos videojogos,
utilizam-se dois computadores ligados em rede: um para o registro do EEG e outro para a execução das atividades
os dois computadores são sincronizados, de modo que o EEG registrado é referenciado aos diversos eventos para cada um dos vídeos jogos;
selecionam-se os trechos do EEG associados a cada um dos eventos de cada jogo.
calcula-se para cada evento da atividade e para cada grupo experimento, a média da atividade elétrica relacionada a esse evento (ARE);
obtém-se a correlação linear da atividade promediada para cada derivação dos AREs em relação à atividade promediada de todas as outras 19 derivações;
calcula-se então a capacidade computacional (h(c)), ou seja, a entropia alocada em cada área ou derivação do EEG para a execução do vídeo jogo, e finalmente
constrói-se os mapas cognitivos cerebrais ( MCCs ) mostrando a capacidade computacional (h(c)) alocada em cada derivação para a execução de um evento da tarefa estudada.
Usou-se, também, análise fatorial para estudar as possíveis correlações de variança entre os valores de h(c) calculados para as distintas derivações do EEG. Dois fatores foram extraidos usando a técnica de Componente Principais, e esses fatores foram rotacionados usando-se o método Varimax Normalizado. Se esses dois fatores explicassem mais de 50% da variabilidade total da h(c), então a análise era considerada válida, e mapas cerebrais eram construídos (chamados aqui de MFs – figs.) codificando em cores os valores do coeficiente de correlação obtido para cada uma das derivações do EEG.
Resultados.
O início da apresentação de cada trecho da história do Currupira (fig. 5) aos surdos recrutou neurônios das áreas centrais (CZ, C3, C4) e frontais (F3, FP2, F4) de ambos os hemisférios, e da área parietal medial (PZ). Após dois segundos de apresentação da história observa-se o envolvimento predominante de neurônios na região frontal e parietal do hemisfério esquerdo (F3, P3), na região frontal e central medial (FZ, CZ), e na região temporal direita (T6). Após quatro segundos o recrutamento envolve áreas bilaterais frontais (F7, F8), áreas mediais da região parietal e central (PZ, CZ), e ainda a área frontal direita (F4). A conectividade média calculada para todo o período de sinalização, mostrou envolvimento de ampla área frontal (F3, F4, FZ, F8), parietal (PZ, P3, P4), e central (C3, C4, CZ), além de OZ. Dois segundos antes do termino da sinalização as mesmas áreas estão ativadas com exceção de FZ, P3 e F8, que deixam de serem recrutadas, e de T6 que passa a ser ativada. Dois segundos após a sinalização as únicas áreas não ativadas são O2, T5, T6, T4, T3, P4 e FP1.
No jogo de charada (fig. 6) foi adicionado mais um evento: A – o momento de escolha da resposta correta. O início da apresentação da informação sinalizada recrutou células das áreas centrais do cérebro (CZ, C3, C4), frontais (F3), com um predomínio do hemisfério direito (F4, F8, FP2), e parietais (PZ). Após dois segundos de sinalização, observou-se uma maior participação de neurônios de ambos os hemisférios nas áreas frontais (F3, F4, F8, F7,FP2), além de neurônios temporais e occipitais direitos (T4, T6, O2). Após quatro segundos a ativação bilateral permanece nas áreas frontais, nas regiões dos eletrodos F3 e F4, e passa também a recrutar bilateralmente as áreas occipitais (O1, O2) e temporais (T5, T6), no hemisfério esquerdo há ainda a ativação de neurônios temporais e centrais (T3, C3), na região medial ocorre ativação de CZ.
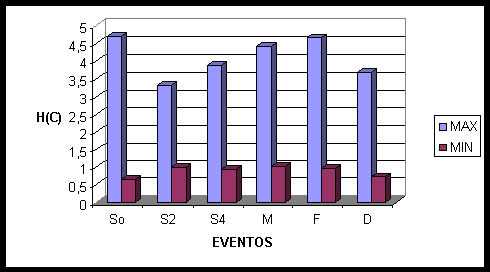
Aqui e na figura seguinte a primeira linha representa os Mapas Cognitivos Cerebrais (MCCs); a segunda e terceira linha representam o resultado das correlações calculadas pela Analise Fatorial, através do Fator 1 e do Fator 2, respectivamente. Os eventos escolhidos são marcados por: 0, 2 e 4 – início da sinalização, dois e quatro segundos após o início; M – média geral de todo o período da sinalização; F – dois segundos antes do término da sinalização; e D – dois segundos após o término da sinalização. O gráfico acima ilustra a entropia mínima e máxima de cada um dos eventos escolhidos.
Na conectividade média aparece o recrutamento de neurônios de todas as áreas cerebrais: occipitais (OZ, O1), temporais (T5), parietais (P3, PZ), centrais (CZ, C3, C4) e frontais (F3, FP2), podemos observar o predomínio de áreas esquerdas (O1, T5, P3, C3, F3). Dois segundos antes do termino da sinalização são recrutados os neurônios da área frontal bilateral (F3, F4), da área central medial (CZ), das áreas occipitais (OZ, O2) e das áreas frontais direitas (F8, FP2). Dois segundos após terminada a sinalização temos uma ativação predominantemente do hemisfério direito, envolvendo áreas frontais (FP2, F4, F8), centrais (C4), e temporais (T4), enquanto no hemisfério esquerdo ocorre ativação somente na área frontal F3. Na região medial temos ativação da área frontal (FZ), central (CZ) e parietal (PZ). No momento do acerto as áreas recrutadas são praticamente todas frontais (F7, F8, F4, FP2, FZ), com exceção de CZ.
Na conectividade média aparece o recrutamento de neurônios de todas as áreas cerebrais: occipitais (OZ, O1), temporais (T5), parietais (P3, PZ), centrais (CZ, C3, C4) e frontais (F3, FP2), podemos observar o predomínio de áreas esquerdas (O1, T5, P3, C3, F3). Dois segundos antes do termino da sinalização são recrutados os neurônios da área frontal bilateral (F3, F4), da área central medial (CZ), das áreas occipitais (OZ, O2) e das áreas frontais direitas (F8, FP2). Dois segundos após terminada a sinalização temos uma ativação predominantemente do hemisfério direito, envolvendo áreas frontais (FP2, F4, F8), centrais (C4), e temporais (T4), enquanto no hemisfério esquerdo ocorre ativação somente na área frontal F3. Na região medial temos ativação da área frontal (FZ), central (CZ) e parietal (PZ). No momento do acerto as áreas recrutadas são praticamente todas frontais (F7, F8, F4, FP2, FZ), com exceção de CZ.
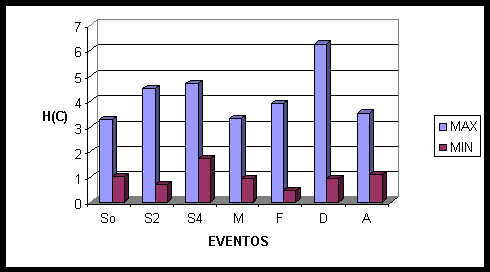
Além da análise baseada no cálculo de entropia das Atividades Relacionadas a Eventos, fizemos uma análise de estatística fatorial utilizando dois fatores, os quais explicam 60% das correlações. O Fator 1 na história do Currupira associa fundamentalmente áreas centrais e frontais, as quais corresponderiam à análise verbal da informação lingüística, enquanto o Fator 2 associa áreas occipitais e áreas temporais e frontais do hemisfério direito, que devem corresponder ao processamento visual gerado pela informação lingüística. No jogo de charada o Fator 1 explica cerca de 50% de toda a variância, mostrando também áreas centrais e frontais correlacionadas entre si.
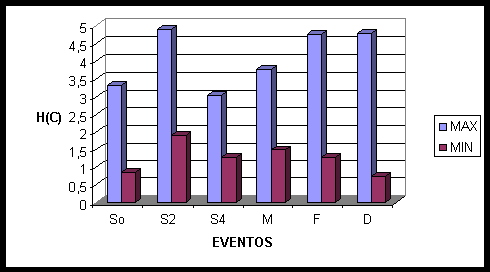
Nos falantes, durante o início da história (fig. 7) temos o recrutamento das áreas mediais parietais e centrais (PZ, CZ), das áreas frontais bilaterais (F7, F8, F4, FP1), além da área central direita (C4). Após dois segundos temos o recrutamento praticamente de áreas frontais mediais e do hemisfério direito (FZ, FP2, F4, F8), além de T4, que se encontra na região temporal direita. Após quatro segundos os voluntários passam a recrutar os neurônios frontais bilaterais e mediais (FP1, FZ, FP2), continuam a ativar T4, e voltam a recrutar a área central medial (CZ). Na conectividade média o recrutamento envolve todas as áreas mediais (OZ, PZ, CZ, FZ), quase toda a área frontal (F7, F8, F4, FP1, FP2), áreas temporais direitas e esquerdas (T6, T4, T3), além da região parietal direita (P4). Dois segundos antes do término da sinalização ocorre unicamente ativação frontal do hemisfério direito (FP2, F4, F8). Dois segundos após a sinalização essas áreas permanecem ativadas, mas passam a se associarem também com áreas mediais (FZ, CZ), frontais esquerdas (F7) e centrais direitas (C4). No acerto ocorre o recrutamento principalmente do hemisfério direito, das áreas frontais (FP2, F4, F8) e centrais (C4), além de F7 e da zona medial (FZ, CZ, PZ).
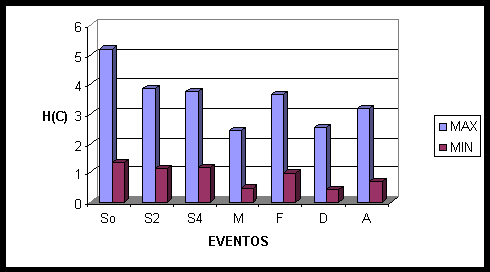
Na charada (fig. 8) os falantes recrutam somente as áreas frontais (F7, FP1, FZ, FP2). Após dois segundos ocorre a associação de neurônios das áreas frontais (F3, F4, F7) com áreas centrais (CZ, C3, C4), e com a área parietal medial (PZ). Após quatro segundos temos a ativação dos eletrodos das áreas frontais mais anteriores (FP1, FZ, FP2, F3), enquanto a outras áreas permanecem as mesmas: centrais (CZ, C3, C4) e parietal medial (PZ). Na conectividade média ocorre o recrutamento de todas as áreas centrais (CZ, C3, C4) e parietais (PZ, P3, P4), de amplas áreas frontais (F7, F3, F4, F8, FP2), e ainda da região occipital medial (OZ). Dois segundos antes de terminar a sinalização houve o recrutamento de neurônios de toda a zona medial (FZ, CZ, PZ, OZ), de áreas frontais direitas (F4, F8) e de áreas frontais e centrais do hemisfério esquerdo (F7, C3). Dois segundos após o término da sinalização as áreas relacionadas são predominantemente do hemisfério esquerdo: frontal (F7), central (C3), temporal (T3, T5), parietal (P3) e occipital (O1), porém ainda ocorre a ativação de OZ, PZ e O1. No acerto são recrutadas as áreas parietais (PZ, P3, P4), centrais (CZ, C3) e frontais (F3, F4), e ainda ocorre ativação das áreas frontais anteriores (FP1, FZ, FP2) e da área temporal esquerda (T3).
Discussão
O fato mais marcante na ativação cerebral dos surdos, observada no presente trabalho, é a associação de neurônios da área parietal medial (PZ) com as áreas centrais (C3, C4) e frontais (F3, F4) bilaterais (fig. 6 e 7), ativação essa que em geral não se observa no processamento cerebral dos falantes, durante tarefas de processamento verbal, como pôde ser constatado em trabalho anterior de Rocha et al, (2000). Como mostrou Neville et al. (1997), dependendo do período de aprendizagem de uma língua, determinadas redes neurais irão se formar de acordo com o estímulo sensorial recebido. Dessa forma, pode-se propor aqui que crianças com surdez congênita, por lhes faltar o estímulo auditivo, tendem a organizar fibras neurais que associem a informação visual ao processamento lingüístico, relacionando áreas visuais do hemisfério direito às áreas lingüísticas do hemisfério esquerdo através da região parietal medial.
Os presentes resultados (fig. 5 e 6) mostram que as áreas verbais frontais (F7, FP1 e F3) e do lóbulo temporal esquerdo (T3 e T5) continuam a trabalhar para o processamento lingüístico, mas agora associadas a neurônios das áreas de decodificação de estímulos visuais e não sonoros. Os estímulos visuais provenientes da linguagem de sinais envolvem expressões faciais, formas e movimentos das mãos, e referência espacial, ou seja, posição das mãos em relação ao resto do corpo. Há no cérebro áreas específicas para o processamento de tais estímulos, uma vez que o manuseio das mãos e dos braços é extremamente importante para todas as atividades que exercemos, e expressões faciais são muito importantes para nosso relacionamento em grupo. Encontra-se ativada nos surdos a área central e parietal esquerda, que justamente contém neurônios que controlam o formato das mãos (Corina et al, 1999), e próxima a ela encontra-se a área de Wernicke, responsável pelo levantamento de informações acerca da semântica de um nome. Além disso a área parietal da mão estabelece conexões com o córtex premotor e provavelmente também com a área de Broca. A posição da mão em relação ao corpo é também identificada por áreas parietais, enquanto o movimento delas é processado por áreas temporais mediais. Áreas de processamento visual de faces localizam-se nas regiões central e parietal do hemisfério direito, as quais também se encontram ativadas pelos surdos durante a visualização e decodificação da Libras. Dessa forma supõe-se que todas essas áreas, relativas ao processamento das mãos e da face, relacionam-se com a área verbal de Broca, com a área nominal de Wernicke e com áreas frontais de memória executiva verbal.
Alguns estudos feitos sobre a linguagem americana de sinais descobriram ativações cerebrais semelhantes àquelas ocorrentes nas línguas faladas (Hickok et al, 1999; Corina et al, 1999; Söderfeldt et al, 1997). No entanto, uma vez que o estímulo sensorial da linguagem de sinais é visual e não auditivo, outras áreas também são ativadas para a decodificação de tal estímulo (Neville et al, 1997; Söderfeldt et al, 1997). Áreas visuais do cérebro também são ativadas pela linguagem durante a comunicação oral entre indivíduos falantes (Rocha et al 2001), porém essa ativação é gerada a partir da imaginação fornecida pela linguagem falada no processo de criar uma imagem visual mental do que é falado. No caso dos surdos essa ativação faz parte do processo de decodificação da própria linguagem além de participar da criação da imagem visual do que é sinalizado. Dessa forma pode-se constatar, a partir dos resultados aqui apresentados, que os surdos apresentam ativações nas áreas referentes ao processamento visual durante a visualização da sinalização da Libras, mas que essa ativação difere da maneira como ela se associa com as áreas verbais do hemisfério esquerdo. Os falantes apresentaram no trabalho de Rocha um maior recrutamento de neurônios das regiões occipitais, associadas às regiões frontais, quando eram submetidos a ouvir uma história e induzidos a imaginá-la visualmente. Aqui os surdos apresentaram associações de áreas frontais, centrais e parietais bilateralmente.
Discussão
O fato mais marcante na ativação cerebral dos surdos, observada no presente trabalho, é a associação de neurônios da área parietal medial (PZ) com as áreas centrais (C3, C4) e frontais (F3, F4) bilaterais (fig. 6 e 7), ativação essa que em geral não se observa no processamento cerebral dos falantes, durante tarefas de processamento verbal, como pôde ser constatado em trabalho anterior de Rocha et al, (2000). Como mostrou Neville et al. (1997), dependendo do período de aprendizagem de uma língua, determinadas redes neurais irão se formar de acordo com o estímulo sensorial recebido. Dessa forma, pode-se propor aqui que crianças com surdez congênita, por lhes faltar o estímulo auditivo, tendem a organizar fibras neurais que associem a informação visual ao processamento lingüístico, relacionando áreas visuais do hemisfério direito às áreas lingüísticas do hemisfério esquerdo através da região parietal medial.
Os presentes resultados (fig. 5 e 6) mostram que as áreas verbais frontais (F7, FP1 e F3) e do lóbulo temporal esquerdo (T3 e T5) continuam a trabalhar para o processamento lingüístico, mas agora associadas a neurônios das áreas de decodificação de estímulos visuais e não sonoros. Os estímulos visuais provenientes da linguagem de sinais envolvem expressões faciais, formas e movimentos das mãos, e referência espacial, ou seja, posição das mãos em relação ao resto do corpo. Há no cérebro áreas específicas para o processamento de tais estímulos, uma vez que o manuseio das mãos e dos braços é extremamente importante para todas as atividades que exercemos, e expressões faciais são muito importantes para nosso relacionamento em grupo. Encontra-se ativada nos surdos a área central e parietal esquerda, que justamente contém neurônios que controlam o formato das mãos (Corina et al, 1999), e próxima a ela encontra-se a área de Wernicke, responsável pelo levantamento de informações acerca da semântica de um nome. Além disso a área parietal da mão estabelece conexões com o córtex premotor e provavelmente também com a área de Broca. A posição da mão em relação ao corpo é também identificada por áreas parietais, enquanto o movimento delas é processado por áreas temporais mediais. Áreas de processamento visual de faces localizam-se nas regiões central e parietal do hemisfério direito, as quais também se encontram ativadas pelos surdos durante a visualização e decodificação da Libras. Dessa forma supõe-se que todas essas áreas, relativas ao processamento das mãos e da face, relacionam-se com a área verbal de Broca, com a área nominal de Wernicke e com áreas frontais de memória executiva verbal.
Alguns estudos feitos sobre a linguagem americana de sinais descobriram ativações cerebrais semelhantes àquelas ocorrentes nas línguas faladas (Hickok et al, 1999; Corina et al, 1999; Söderfeldt et al, 1997). No entanto, uma vez que o estímulo sensorial da linguagem de sinais é visual e não auditivo, outras áreas também são ativadas para a decodificação de tal estímulo (Neville et al, 1997; Söderfeldt et al, 1997). Áreas visuais do cérebro também são ativadas pela linguagem durante a comunicação oral entre indivíduos falantes (Rocha et al 2001), porém essa ativação é gerada a partir da imaginação fornecida pela linguagem falada no processo de criar uma imagem visual mental do que é falado. No caso dos surdos essa ativação faz parte do processo de decodificação da própria linguagem além de participar da criação da imagem visual do que é sinalizado. Dessa forma pode-se constatar, a partir dos resultados aqui apresentados, que os surdos apresentam ativações nas áreas referentes ao processamento visual durante a visualização da sinalização da Libras, mas que essa ativação difere da maneira como ela se associa com as áreas verbais do hemisfério esquerdo. Os falantes apresentaram no trabalho de Rocha um maior recrutamento de neurônios das regiões occipitais, associadas às regiões frontais, quando eram submetidos a ouvir uma história e induzidos a imaginá-la visualmente. Aqui os surdos apresentaram associações de áreas frontais, centrais e parietais bilateralmente.
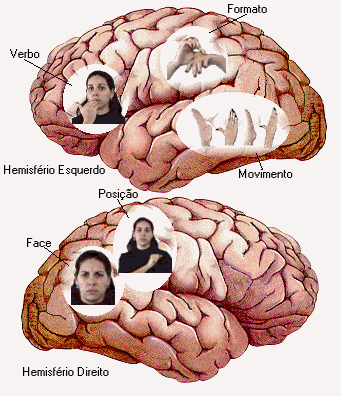
Fig. 9 Processamento lingüístico da Libras
Na Fig. 9, à direita, observa-se que as áreas responsáveis pela decodificação do movimento, das formas e das posições das mãos, além da área de identificação de faces, se encontram distribuídas ao longo de toda a região central e parietal de ambos os hemisférios. Essas áreas, para os surdos, fornecem a informação necessária para Broca e Wernicke definirem as ações e os objetos das palavras descritas pela língua de sinais, através de gestos motores, da mesma maneira como as áreas de Broca e de Wernicke, no caso da língua falada, definem a semântica e a sintaxe do verbo comer e a semântica de seus complementos.
Observando a analise fatorial pode-se constatar que o Fator 1 agrupou áreas centrais e frontais do hemisfério esquerdo (F3, C3 – áreas verbais) e direito (F4, C4 – áreas visuais), enquanto o Fator 2 mostra um maior agrupamento de neurônios occipitais relacionados ao processamento visual. Pode-se hipotetizar dessa forma que a informação visual é decodificada nas áreas occipitais quando não faz parte do processamento lingüístico, enquanto que ao pertencer à fonologia ou morfologia da língua de sinais passa a ser processada pelas áreas visuais mais anteriores (F4, C4), adjacentes às áreas de processamento lingüístico (F3, C3).
Nos falantes, que tiveram um aprendizado tardio da Libras, nota-se uma maior ativação de áreas frontais, mediais e temporais. A Libras, por se tratar de uma segunda língua, parece exigir dos falantes uma tradução dos sinais para a língua portuguesa, associando dessa forma áreas frontais bilaterais de memória verbal e visual com áreas temporais de processamento lingüístico. Essa diferença no padrão de ativação cerebral entre surdos e falantes mostra a necessidade de se aprender uma língua de sinais na infância, para que ela seja diretamente decodificada pelo seu próprio padrão neural (Neville et al, 1997).
De todos os métodos de comunicação propostos para os surdos a língua de sinais foi a que mais deu certo, e isso se comprova cientificamente pela facilidade natural do homem de associar áreas cerebrais responsáveis por todas as informações visuais veiculadas pela Libras às áreas cerebrais responsáveis pela significação dos nomes (Wernicke), das ações (Broca) e das sentenças (área frontal esquerda). A língua de sinais deve ser para o surdo uma língua materna que possa funcionar de ferramenta para o aprendizado de uma segunda língua também visual: a escrita das línguas faladas. Assim a melhor maneira de englobar os surdos à sociedade majoritária é fornecer-lhes uma maneira fácil e natural de se comunicarem entre si e de aprenderem como se comunicarem com os ouvintes, através de um ensino precoce da Libras e do português em sua forma visual (leitura) e motora (escrita).
Os resultados obtidos nessa pesquisa ajudam a compreender o funcionamento cerebral da Libras, e a comprovar que se trata de uma linguagem tão complexa e sofisticada como qualquer outra. No Brasil seu uso ainda se encontra mal difundido, e sua compreensão mal estudada. Torna-se necessário uma maior atenção tanto por parte dos pesquisadores quanto por parte dos educadores, para tornar a língua de sinais uma língua fluente entre aqueles que a necessitam e aqueles que convivem com surdos. Mesmo em países com uma maior estrutura para ensino da língua de sinais, seu conhecimento neurológico se encontra pouco estudado, porém seu uso já é muito mais difundido criando com isso toda uma cultura para a língua e seus sinalizadores. No Brasil podemos caminhar simultaneamente para o progresso do ensino e do conhecimento da Libras e do desenvolvimento de sua cultura.
Na Fig. 9, à direita, observa-se que as áreas responsáveis pela decodificação do movimento, das formas e das posições das mãos, além da área de identificação de faces, se encontram distribuídas ao longo de toda a região central e parietal de ambos os hemisférios. Essas áreas, para os surdos, fornecem a informação necessária para Broca e Wernicke definirem as ações e os objetos das palavras descritas pela língua de sinais, através de gestos motores, da mesma maneira como as áreas de Broca e de Wernicke, no caso da língua falada, definem a semântica e a sintaxe do verbo comer e a semântica de seus complementos.
Observando a analise fatorial pode-se constatar que o Fator 1 agrupou áreas centrais e frontais do hemisfério esquerdo (F3, C3 – áreas verbais) e direito (F4, C4 – áreas visuais), enquanto o Fator 2 mostra um maior agrupamento de neurônios occipitais relacionados ao processamento visual. Pode-se hipotetizar dessa forma que a informação visual é decodificada nas áreas occipitais quando não faz parte do processamento lingüístico, enquanto que ao pertencer à fonologia ou morfologia da língua de sinais passa a ser processada pelas áreas visuais mais anteriores (F4, C4), adjacentes às áreas de processamento lingüístico (F3, C3).
Nos falantes, que tiveram um aprendizado tardio da Libras, nota-se uma maior ativação de áreas frontais, mediais e temporais. A Libras, por se tratar de uma segunda língua, parece exigir dos falantes uma tradução dos sinais para a língua portuguesa, associando dessa forma áreas frontais bilaterais de memória verbal e visual com áreas temporais de processamento lingüístico. Essa diferença no padrão de ativação cerebral entre surdos e falantes mostra a necessidade de se aprender uma língua de sinais na infância, para que ela seja diretamente decodificada pelo seu próprio padrão neural (Neville et al, 1997).
De todos os métodos de comunicação propostos para os surdos a língua de sinais foi a que mais deu certo, e isso se comprova cientificamente pela facilidade natural do homem de associar áreas cerebrais responsáveis por todas as informações visuais veiculadas pela Libras às áreas cerebrais responsáveis pela significação dos nomes (Wernicke), das ações (Broca) e das sentenças (área frontal esquerda). A língua de sinais deve ser para o surdo uma língua materna que possa funcionar de ferramenta para o aprendizado de uma segunda língua também visual: a escrita das línguas faladas. Assim a melhor maneira de englobar os surdos à sociedade majoritária é fornecer-lhes uma maneira fácil e natural de se comunicarem entre si e de aprenderem como se comunicarem com os ouvintes, através de um ensino precoce da Libras e do português em sua forma visual (leitura) e motora (escrita).
Os resultados obtidos nessa pesquisa ajudam a compreender o funcionamento cerebral da Libras, e a comprovar que se trata de uma linguagem tão complexa e sofisticada como qualquer outra. No Brasil seu uso ainda se encontra mal difundido, e sua compreensão mal estudada. Torna-se necessário uma maior atenção tanto por parte dos pesquisadores quanto por parte dos educadores, para tornar a língua de sinais uma língua fluente entre aqueles que a necessitam e aqueles que convivem com surdos. Mesmo em países com uma maior estrutura para ensino da língua de sinais, seu conhecimento neurológico se encontra pouco estudado, porém seu uso já é muito mais difundido criando com isso toda uma cultura para a língua e seus sinalizadores. No Brasil podemos caminhar simultaneamente para o progresso do ensino e do conhecimento da Libras e do desenvolvimento de sua cultura.
Fonte:
http://www.enscer.com.br/pesquisas/artigos/libras/libras.html
Nenhum comentário:
Postar um comentário
COMENTE AQUI NO BLOG!!!
SEU COMENTÁRIO FAZ TODA DIFERENÇA!!!
Um comentário é o que você pensa, sua opinião, alguma coisa que você quer falar comigo.
BJOS SINALIZADOS.